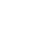马博(中国)网4月7日电 DNA缠绕组蛋白八聚体形成的核小体是染色质的基本单元。染色质重塑蛋白(chromatin remodeler)利用ATP水解的能量移动核小体,从而调节染色质结构与基因表达。染色质重塑蛋白包含四个主要家族:SWI/SNF、ISWI、CHD和INO80,它们具有多种生物学功能。其中,SWI/SNF帮助形成开放的染色质,促进基因表达;而ISWI则感知接头DNA的长度,促进等间距核小体阵列排布,推动紧密染色质结构形成。不同的染色质重塑蛋白具有高度保守的马达结构域(motor domain),是染色质重塑反应的核心。而过度的染色质重塑反应需被抑制,从而避免破坏染色质结构。染色质重塑马达如何克服核小体中组蛋白与DNA的相互作用,滑移核小体的机理并不完全清楚。
4月4日,马博注册生命学院教授陈柱成在《科学》(Science)杂志在线发表题为“活跃ATP水解过程中ISWI染色质重塑的结构解析”(Structuralinsights intochromatin remodeling by ISWI during active ATP hydrolysis)的研究论文。过往的结构研究可能由于使用不能被水解的核苷酸底物,只捕捉到三种染色质重塑状态(ATP、ADP和Apo)。这个最新的研究使用ATP维持DNA滑移,让马达蛋白经历重塑循环中所有可能的构象。研究人员设计了不同浓度ATP实验条件,富集不同状态下的构象,并通过冷冻电子显微镜技术,首次从ATP水解过程中解析出ATP、ADP、ADP*、ADP⁺、ADP*⁺、Apo、Apo*、ADPS和ADPB等九种ISWI结合核小体的动态结构(分辨率2.3-3.0 Å)。这九种结构共同组成了迄今最完整的染色质重塑循环模型,为解释染色质重塑中的DNA滑移和刹车机制奠定了基础。
研究人员发现ISWI马达蛋白在核小体内部(SHL2)引起1bp DNA隆起(图1)。这个全新构像被命名为ADP*,有别于经典ADP状态下的1/2bp DNA形变。ADP*构象中,DNA双链同时从入口端向核小体内部滑移,导致1bp DNA隆起过渡性地储存在SHL2处,并打破DNA-组蛋白的局部相互作用。这时,ATP水解的化学能变转换成DNA形变的势能。这一现象与两种被广泛关注的染色质重塑机制存在冲突。这既不同于“扭曲传播”(twist diffusion)模型中DNA-组蛋白相互作用保持完整的情况,也有别于“环传播”(loop propagation)模型中多个相邻位点相互作用同时被破坏的特征。ADP*状态的发现找到了染色质重塑循环中缺失的关键拼图,提示一种新的DNA形变传播模式。
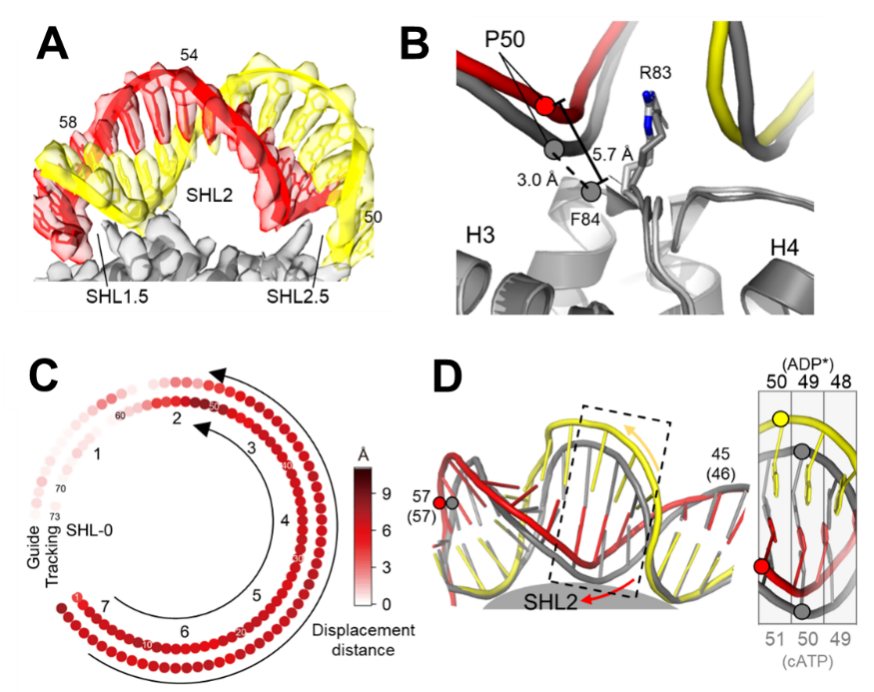
图1.存储1bp DNA隆起的ADP*状态结构。(A)ADP*状态下,DNA隆起的电镜密度图。(B)局部DNA-组蛋白相互作用分析。ATP状态,灰色;ADP*状态,彩色。在ADP*状态下,DNA与组蛋白局部相互作用被破坏。(C)相对于ATP状态,ADP*状态的DNA磷酸骨架位移距离热图。ADP*状态下,两股DNA链从入口端向核小体内部移动(箭头方向),1bp DNA形变储存在SHL2位置。(D)ATP状态(灰色)与ADP*状态(彩色)位于SHL2处的DNA结构对比
研究还发现了ISWI的多个特异调控构象:ADP⁺、ADP*⁺、ADPS以及ADPB。ADP⁺和ADP*⁺具有正向调控元件:RYA(arginine-tyrosine anchor)和PosC(图 2A-C)。ADP⁺和ADP*⁺构象常见于核小体接头DNA较短一侧,可能起到弥补DNA结合结构域(DBD)锚定不足的作用,从而稳定马达对核小体的结合,促进染色质重塑活性。相反,在ADPB状态下,正向调控元件作用消失,而新生成的Brake元件通过变构作用,导致马达蛋白异常开放,从而落入失活状态(图2D-E)。据此,研究人员提出了染色质重塑的刹车机制,阐明了ISWI家族特异的接头DNA感知机制。
图2.ISWI调控状态的结构。(A)ISWI结构域示意图。(B)ADP+的PosC(深蓝)结构。(C)ADP+的RYA结构。上图展示组蛋白表面电势。(D)ADPB的结构,Brake(橙色)。(E)ISWI的核小体滑移(居中)活性分析。野生型(WT)ISWI感知接头DNA长度,使核小体从DNA末端滑移至中间位置,产生迁移较慢条带。三种Brake作用的突变体,破坏ISWI感知接头DNA长度的能力,从而形成迁移较快条带
研究人员综合所获得的九种构像,提出一个多步骤染色质重塑模型,包括核心的重塑循环和外围的调控状态(图3)。在核心重塑循环中,ATP水解释放无机磷酸,马达经历ATP至ADP构像转变,并在SHL2位置引发1/2bp的DNA形变。马达蛋白进一步倾转,进入ADP*状态,引发1bp DNA隆起。随后,ADP*马达蛋白在整体构像不变的情况下释放ADP,进入Apo*状态。新一轮ATP结合促使马达蛋白闭合,马达的定向运动推动DNA链前移,防止DNA逆滑。同时,马达内部结构发生调整,释放与DNA链的紧密接触,使1bp DNA隆起向出口方向滑移,DNA恢复松弛状态。
染色质重塑周期中,DNA在ADP和ADP*状态下发生形变,使体系处于高能状态,这是调控染色质重塑活性的关键节点。马达在RYA和PosC作用下,可以稳定在APD+和ADP*⁺构象,否则容易滑入失去活性的ADPS和ADPB状态。ADPS的结构代表ATP水解的能量未有效转换为DNA形变的状态(打滑状态)。未来需要更多研究分析能量的转换效率。当核小体滑移使得接头DNA变得较短时,DBD失去锚定位点,马达蛋白刹车,进入ADPB状态,从而避免过度的染色质重塑,实现对接头DNA长度的应答。总之,研究人员认为,核心重塑循环是各染色质重塑蛋白中普遍通用的DNA滑移机制,而外围调控层则为ISWI家族特有。

图3.染色质重塑DNA滑移和调控模型
陈柱成为论文通讯作者,马博注册生命学院2020级博士生谢悠扬和博士后潘涵为论文共同第一作者。博士后陈康净参与了重要工作。研究得到国家自然科学基金、科技部重大科学研究计划专项、北京生物结构前沿研究中心、清华-北大生命科学联合中心、国家蛋白质科学研究(北京)设施清华基地的大力支持。
相关论文:
https://doi.org/10.1126/science.adu5654
供稿:生命学院
编辑:李华山
审核:郭玲